Patagium and tail morphology shape aerodynamic performance and control authority in gliding-mammal-inspired wings
Pith reviewed 2026-07-03 04:54 UTC · model grok-4.3
The pith
Gliding-mammal-inspired wing morphologies create distinct aerodynamic trade-offs in lift, drag, and control authority.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
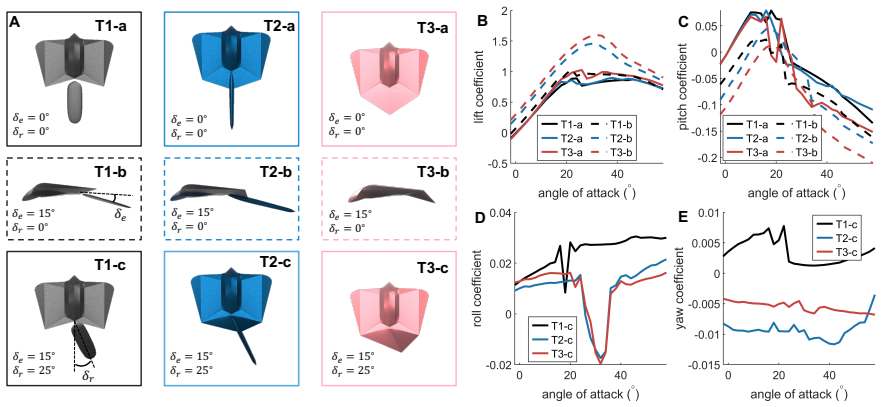
Computational fluid dynamics simulations of three patagium configurations showed that a broader outline produced the highest lift and lift coefficient while an intermediate morphology provided a smoother post-stall response with lower drag. For tail configurations, the integrated uropatagium improved lift and pitch control under symmetric deflection, whereas the flat tail generated stronger rolling and yawing responses under asymmetric deflection. These results establish that gliding-mammal-inspired morphologies produce distinct aerodynamic trade-offs rather than a single optimal design.
What carries the argument
CFD comparisons that isolate patagium membrane outline and tail/uropatagium geometry under baseline, symmetric-deflection, and asymmetric-deflection conditions to quantify forces and moments.
If this is right
- A broader patagium outline maximizes lift and lift coefficient.
- An intermediate patagium outline reduces drag and smooths post-stall behavior.
- A colugo-like integrated uropatagium increases lift and pitch-control authority under symmetric deflection.
- A flat-tail configuration produces stronger rolling and yawing responses under asymmetric deflection.
- No single morphology optimizes all aerodynamic and control metrics simultaneously.
Where Pith is reading between the lines
- The observed trade-offs may explain why different gliding mammal species evolved distinct patagium and tail shapes suited to their typical environments or behaviors.
- Morphing aerial robots could switch between patagium and tail shapes mid-flight to prioritize lift during climb versus lateral control during turns.
- Free-flight wind-tunnel or outdoor tests of the same shapes would reveal whether the fixed-condition CFD results hold when the wing is allowed to move and respond to its own wake.
- Similar morphology-performance mapping could be applied to other gliding animals such as flying squirrels to test whether the same trade-off patterns appear.
Load-bearing premise
The chosen representative morphologies and controlled CFD flow conditions are representative enough of real gliding mammal flight to isolate the separate effects of patagium and tail on performance and control.
What would settle it
Force and moment measurements on physical gliding-mammal models or live animals executing comparable maneuvers that show different relative lift coefficients, drag values, or control moments than the CFD results.
Figures





read the original abstract
Gliding mammals exhibit diverse patagium and tail/uropatagium morphologies that may influence aerodynamic performance and maneuverability. Here, we use computational fluid dynamics to isolate the aerodynamic effects of representative gliding-mammal-inspired morphologies under controlled flow conditions. Three patagium configurations were compared to evaluate the effects of membrane outline on lift generation, drag, stall behavior and pitching moment. Three tail/uropatagium configurations were further tested under baseline, symmetric-deflection and asymmetric-deflection conditions to assess their longitudinal and lateral control authority. The results show that a broader patagium configuration generated the highest lift and lift coefficient, whereas an intermediate patagium morphology showed a smoother post-stall response with lower drag. For the tail configurations, the colugo-like integrated uropatagium enhanced lift and pitch-control authority under symmetric deflection, while the flat-tail configuration produced stronger rolling and yawing responses under asymmetric deflection. These findings indicate that gliding-mammal-inspired morphologies produce distinct aerodynamic trade-offs rather than a single optimal design. The results provide insight into the functional diversity of gliding mammal morphology and offer design guidance for bioinspired morphing aerial robots.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses computational fluid dynamics to isolate the effects of three representative patagium outlines and three tail/uropatagium shapes on lift, drag, stall behavior, and pitching/rolling/yawing moments under controlled flow conditions. It reports that a broader patagium produces the highest lift coefficient, an intermediate patagium yields smoother post-stall response with lower drag, a colugo-like integrated uropatagium enhances lift and pitch-control authority under symmetric deflection, and a flat-tail configuration produces stronger rolling and yawing responses under asymmetric deflection. The central claim is that these morphologies generate distinct aerodynamic trade-offs rather than a single optimal design, providing insight into gliding-mammal functional diversity and guidance for bioinspired morphing aerial robots.
Significance. If the CFD results are reliable, the work supplies concrete, morphology-specific trade-offs (e.g., lift vs. post-stall smoothness, pitch vs. roll/yaw authority) that advance understanding of why gliding mammals exhibit diverse patagium and tail forms and that can directly inform parameter choices in morphing-wing robot design.
major comments (1)
- [Methods] Methods section (and abstract): no Reynolds-number values, turbulence-model specification, mesh-convergence data, or validation against experiments, wind-tunnel benchmarks, or animal flight data are supplied. This is load-bearing because the reported differences in lift coefficients, stall behavior, and control moments rest entirely on the fidelity of the simulated flow fields under the chosen conditions.
minor comments (1)
- [Abstract] Abstract: quantitative deltas (e.g., percentage lift increase or moment coefficient changes) between configurations would strengthen the comparative claims.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback. The single major comment identifies a clear gap in the methods documentation that we will address directly in revision.
read point-by-point responses
-
Referee: [Methods] Methods section (and abstract): no Reynolds-number values, turbulence-model specification, mesh-convergence data, or validation against experiments, wind-tunnel benchmarks, or animal flight data are supplied. This is load-bearing because the reported differences in lift coefficients, stall behavior, and control moments rest entirely on the fidelity of the simulated flow fields under the chosen conditions.
Authors: We agree that these details are essential for evaluating the CFD results. In the revised manuscript we will (i) state the Reynolds number(s) based on the reference length and freestream velocity used, (ii) specify the turbulence model and closure constants, (iii) present mesh-convergence data (force coefficients and moment coefficients versus cell count) demonstrating that the reported differences remain within acceptable tolerances, and (iv) add a validation subsection that compares the solver setup against published wind-tunnel data for comparable low-aspect-ratio wings and any available gliding-mammal kinematic or force measurements. We note that comprehensive live-animal validation data remain sparse in the literature; the revised text will therefore frame the validation as a combination of canonical benchmarks and sensitivity checks rather than direct animal replication. revision: yes
Circularity Check
No circularity: results are direct CFD outputs with no fitted predictions or self-referential derivations
full rationale
The paper conducts CFD simulations comparing three patagium outlines and three tail/uropatagium shapes under controlled conditions, reporting lift, drag, stall behavior, and control moments as direct simulation outputs. No equations, parameter fits, or self-citations are described that would reduce reported coefficients to inputs by construction. The central claim of distinct aerodynamic trade-offs follows from comparative simulation results rather than any definitional or fitted equivalence. This is a standard numerical-experiment paper whose load-bearing steps are external to the reported values.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption CFD simulations with the chosen turbulence model and boundary conditions accurately capture the aerodynamic forces and moments on the tested morphologies
Reference graph
Works this paper leans on
-
[1]
Gliding mammals of the world.CSIRP Pub, 156, 2012
S Jackson. Gliding mammals of the world.CSIRP Pub, 156, 2012
2012
-
[2]
Soft biohybrid morphing wings with feathers underactuated by wrist and finger motion.Science Robotics, 5(38):eaay1246, 2020
Eric Chang, Laura Y Matloff, Amanda K Stowers, and David Lentink. Soft biohybrid morphing wings with feathers underactuated by wrist and finger motion.Science Robotics, 5(38):eaay1246, 2020
2020
-
[3]
Bird-inspired reflexive morphing enables rudderless flight.Science Robotics, 9(96):eado4535, 2024
Eric Chang, Diana D Chin, and David Lentink. Bird-inspired reflexive morphing enables rudderless flight.Science Robotics, 9(96):eado4535, 2024
2024
-
[4]
Bioinspired wing and tail morphing extends drone flight capabilities.Science Robotics, 5(47):eabc2897, 2020
Enrico Ajanic, Mir Feroskhan, Stefano Mintchev, Flavio Noca, and Dario Floreano. Bioinspired wing and tail morphing extends drone flight capabilities.Science Robotics, 5(47):eabc2897, 2020
2020
-
[5]
A twist of the tail in turning maneuvers of bird- inspired drones.Science Robotics, 9(96):eado3890, 2024
Hoang-Vu Phan and Dario Floreano. A twist of the tail in turning maneuvers of bird- inspired drones.Science Robotics, 9(96):eado3890, 2024. 12
2024
-
[6]
F. T. Muijres, L. C. Johansson, R. Barfield, M. Wolf, G. R. Spedding, and A. Hedenström. Leading-edge vortex improves lift in slow-flying bats.Science, 319(5867):1250–1253, 2008
2008
-
[7]
Global dynamics of non-equilibrium gliding in animals.Bioinspiration & Biomimetics, 12(2):026013, 2017
Isaac J Yeaton, John J Socha, and Shane D Ross. Global dynamics of non-equilibrium gliding in animals.Bioinspiration & Biomimetics, 12(2):026013, 2017
2017
-
[8]
Monopedal robot branch-to-branch leaping and landing inspired by squirrel balance control.Science Robotics, 10(100):eadq1949, 2025
Justin K Yim, Eric K Wang, Sebastian D Lee, Nathaniel H Hunt, Robert J Full, and Ronald S Fearing. Monopedal robot branch-to-branch leaping and landing inspired by squirrel balance control.Science Robotics, 10(100):eadq1949, 2025
2025
-
[9]
Aerodynamic charac- teristics and pitching adjusting mechanism of the flying squirrel with deployed patagium
Fei Zhao, Wei Wang, Jingtao Zhang, Justyna Wyrwa, and Feng Sun. Aerodynamic charac- teristics and pitching adjusting mechanism of the flying squirrel with deployed patagium. IEEE Access, 7:185554–185564, 2019
2019
-
[10]
A squirrel-inspired drone with enhanced stability, agility and maneuverability via whole-body morphing.Nature Commu- nications, 2026
Liming Zheng, Alexander van Zuijlen, and Salua Hamaza. A squirrel-inspired drone with enhanced stability, agility and maneuverability via whole-body morphing.Nature Commu- nications, 2026
2026
-
[11]
Reproductive biology of the red-giant flying squirrel, petaurista petaurista, in taiwan.Journal of mammalogy, 74(4):982–989, 1993
Pei-Fen Lee, Yao-Sung Lin, and Donald R Progulske. Reproductive biology of the red-giant flying squirrel, petaurista petaurista, in taiwan.Journal of mammalogy, 74(4):982–989, 1993
1993
-
[12]
The relationship between 3-d kinematics and gliding performance in the southern flying squirrel, glaucomys volans.Journal of Experimental Biology, 209(4):689– 701, 2006
Kristin L Bishop. The relationship between 3-d kinematics and gliding performance in the southern flying squirrel, glaucomys volans.Journal of Experimental Biology, 209(4):689– 701, 2006
2006
-
[13]
Greg Byrnes, Norman T-L Lim, and Andrew J Spence. Take-off and landing kinetics of a free-ranging gliding mammal, the malayan colugo (galeopterus variegatus).Proceedings of the Royal Society B: Biological Sciences, 275(1638):1007–1013, 2008
2008
-
[14]
Sex differences in the locomotor ecology of a gliding mammal, the malayan colugo (galeopterus variegatus)
Greg Byrnes, Norman T-L Lim, Charlene Yeong, and Andrew J Spence. Sex differences in the locomotor ecology of a gliding mammal, the malayan colugo (galeopterus variegatus). Journal of Mammalogy, 92(2):444–451, 2011
2011
-
[15]
John Wiley & Sons, 2004
Warren F Phillips.Mechanics of flight. John Wiley & Sons, 2004
2004
-
[16]
Seamless active morphing wing simultaneous gust and maneuver load alleviation.Journal of Guidance, Control, and Dynamics, 44(9):1649–1662, 2021
Xuerui Wang, Tigran Mkhoyan, Iren Mkhoyan, and Roeland De Breuker. Seamless active morphing wing simultaneous gust and maneuver load alleviation.Journal of Guidance, Control, and Dynamics, 44(9):1649–1662, 2021
2021
-
[17]
Glide performance and aerodynamics of non-equilibrium glides in northern flying squirrels (glau- comys sabrinus).Journal of the Royal Society interface, 10(80):20120794, 2013
Joseph W Bahlman, Sharon M Swartz, Daniel K Riskin, and Kenneth S Breuer. Glide performance and aerodynamics of non-equilibrium glides in northern flying squirrels (glau- comys sabrinus).Journal of the Royal Society interface, 10(80):20120794, 2013
2013
-
[18]
Cfd simulation of flow past mav wings.International Journal of Aerospace Innovations, 5(1), 2013
Pradeep Shetty, MB Subrahmanya, DS Kulkarni, and BN Rajani. Cfd simulation of flow past mav wings.International Journal of Aerospace Innovations, 5(1), 2013
2013
-
[19]
Xfoil vs cfd performance predictions for high lift low reynolds number airfoils.Aerospace Science and Technology, 52:207–214, 2016
JCPJ Morgado, R Vizinho, MAR Silvestre, and JC Páscoa. Xfoil vs cfd performance predictions for high lift low reynolds number airfoils.Aerospace Science and Technology, 52:207–214, 2016
2016
-
[20]
Numerical investigation of low-aspect-ratio wings at low reynolds numbers.Journal of Aircraft, 43(3):713–722, 2006
Peter Cosyn and Jan Vierendeels. Numerical investigation of low-aspect-ratio wings at low reynolds numbers.Journal of Aircraft, 43(3):713–722, 2006. 13
2006
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.