Single-cell-level distributions and relationships can differentiate cell-division and growth models
Pith reviewed 2026-07-02 05:36 UTC · model grok-4.3
The pith
Probability distributions of cell sizes and times differentiate Timer, Sizer, Adder models and linear versus exponential growth.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Probability distributions and statistical relationships among birth-size, division-size, division-time, and added-size serve as discriminants for both the division timing rules (Timer, Sizer, Adder) and the growth law (linear or exponential), remaining effective under stochastic single-cell growth, asymmetric partitioning, and correlated growth rates across generations.
What carries the argument
The probability distributions of physiological quantities (birth-size, division-size, division-time, division-added-size) and their inter-relationships under stochastic growth and asymmetric partitioning for the Timer, Sizer, and Adder models.
If this is right
- Distinct distribution shapes arise for each division model under stochastic conditions.
- Joint distributions differ between linear and exponential growth for the same division rule.
- The differentiability holds when growth rates correlate between mother and daughter cells.
- Analytical predictions match experimental single-cell measurements for microbial systems.
Where Pith is reading between the lines
- Measuring these distributions in a new organism could identify its division rule without knowing the molecular network.
- The method could be applied to systems with additional noise sources to test robustness of the signatures.
- Extensions to multi-generational data might reveal higher-order correlations beyond pairwise ones.
Load-bearing premise
The specific mathematical forms chosen for stochastic growth, asymmetric division, and the three division rules capture the dominant statistical features that experiments would observe.
What would settle it
Observing that the measured distributions for cells known to follow the Adder model are statistically indistinguishable from those predicted for the Sizer model would falsify the claim of differentiability.
Figures



read the original abstract
Complex interactions among regulatory molecules determine the rules underlying cell growth and division in microbial cells. While the governing molecular network may not always be obvious, it is well known that correlations among certain physiological quantities measured in experiments, such as birth-size, division-size, division-time, and division-added-size, can differentiate among various cell-division models, such as Timer, Sizer, and Adder. Here we show that, apart from these correlations, which we extend for the case of stochastic single-cell growth and stochastic asymmetric partitioning, probability distributions of these quantities and statistical relationships between them can also be used to differentiate between these division models. Interestingly, we show that these quantities can not only differentiate the division models, but also distinguish among the single-cell growth paradigms, such as linear and exponential growth. We then demonstrate this differentiability among various division and growth models by comparing our analytical results with published experimental data. We further show that these results remain valid even when the growth rate of a cell is correlated with the growth rate of cells from previous generations in the lineage.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that, in addition to correlations, the probability distributions of birth size, division size, division time, and added size, together with statistical relationships among them, can distinguish Timer/Sizer/Adder division rules and linear versus exponential growth even under stochastic single-cell growth, asymmetric partitioning, and lineage-correlated growth rates. Analytical expressions for these distributions are derived and compared to published experimental data to demonstrate the differentiability.
Significance. If the derivations are correct and the reported signatures remain identifiable, the work supplies additional observables beyond mean correlations for model discrimination in microbial cell-cycle studies, which could improve inference of division strategies from single-cell measurements.
major comments (2)
- [Abstract and results sections on analytical derivations] The differentiability is derived under specific functional forms for stochastic growth and partitioning; the manuscript does not test whether the reported signatures remain unique once additional regulatory feedback, different noise spectra, or measurement error are introduced. This robustness question is load-bearing for the claim that the distributions 'can differentiate' models in general experimental settings.
- [Section discussing lineage correlations] The extension to correlated growth rates across generations is shown, but the argument provides no concrete test (e.g., via simulation or perturbation analysis) that the distinguishing power survives unmodeled mechanisms beyond the three canonical rules and two growth paradigms.
Simulated Author's Rebuttal
We thank the referee for the comments on robustness. Our derivations provide exact distributions and relations under the stated stochastic growth, partitioning, and lineage-correlation assumptions for the canonical Timer/Sizer/Adder and linear/exponential models. We address the two major comments below and will make targeted revisions to clarify scope and limitations.
read point-by-point responses
-
Referee: [Abstract and results sections on analytical derivations] The differentiability is derived under specific functional forms for stochastic growth and partitioning; the manuscript does not test whether the reported signatures remain unique once additional regulatory feedback, different noise spectra, or measurement error are introduced. This robustness question is load-bearing for the claim that the distributions 'can differentiate' models in general experimental settings.
Authors: We agree the analysis assumes specific functional forms (e.g., multiplicative noise for growth rates and beta-distributed partitioning). The manuscript derives closed-form distributions and shows they differ across the six model combinations, with validation against published data. We do not claim the signatures are invariant to arbitrary additional feedback, noise spectra, or measurement error. We will revise the abstract and discussion to state the assumptions explicitly and note that robustness to unmodeled mechanisms remains an open question, while the reported signatures already differentiate the canonical cases under the included stochasticity. revision: partial
-
Referee: [Section discussing lineage correlations] The extension to correlated growth rates across generations is shown, but the argument provides no concrete test (e.g., via simulation or perturbation analysis) that the distinguishing power survives unmodeled mechanisms beyond the three canonical rules and two growth paradigms.
Authors: The lineage-correlation extension yields modified analytical expressions that remain distinct across models; we compare these to data. We provide no additional perturbation analysis or simulations for mechanisms outside the three rules and two growth modes. We will add a short discussion paragraph acknowledging this scope limitation and that distinguishing power could be affected by other unmodeled processes. revision: partial
Circularity Check
Derivations start from explicit model assumptions without reduction to self-referential inputs
full rationale
The paper starts from standard functional forms for stochastic growth (linear/exponential), asymmetric partitioning, and the three division rules (Timer/Sizer/Adder), then derives closed-form or semi-analytic expressions for the distributions of birth/division sizes, times, and added sizes plus their correlations. These expressions are obtained directly from the assumed stochastic processes and are compared to external experimental data. No load-bearing step reduces by construction to a fitted parameter renamed as a prediction, a self-citation chain, or an ansatz smuggled from prior work by the same authors. The claimed differentiability is a direct mathematical consequence of the input model definitions, which remain independent of the target distinctions. The extension to lineage-correlated growth rates is likewise obtained by modifying the same stochastic assumptions rather than by re-using the result itself.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Timer model Consider a Mother Machine setup in which individual cells are tracked over multiple generations. We assume that cells grow exponentially in size and divide symmetrically into two equal daughter cells without partitioning stochastic- ity, following the Timer model of cell division. The birth-size for a daughter cell is given bysbd =s b exp (ατd...
-
[2]
The first argument goes like this
Sizer and Adder models For the Sizer and Adder models, we present some statistical arguments that are used to obtainζ(s b) from principal distributions. The first argument goes like this. The birth-size of a random cell in a lineage is given bys b =βs d. Here,s d is the size of the parent cell at the time of division, and is sampled from the probability d...
-
[3]
Probability transformation of random variables Suppose we are given a joint probability distribution of two random variablesxandyasP(x, y). The probability distribution for another variablez=Z(x, y) is given as [61] : Q(z) = Z ∞ 0 Z ∞ 0 P(x, y)δ(z−Z(x, y))dx dy.(B1) Note that the lower limit of integration is 0, not−∞, because the variables we have consid...
-
[4]
A5, as discussed in Appendix A
Sizer model Apart from the free-parameter distributions,ζ(s b) for the Sizer model is obtained from Ξ(s d) andκ(β) using Eq. A5, as discussed in Appendix A. The other single-cell-level distributions for the Sizer model, i.e., Γ(τ d) and Ω(∆ d), can also be obtained from Ξ(s d) andζ(s b) using probability transformations. We already have Ξ(s d) andζ(s b) f...
-
[5]
The other single-cell-level distributions for the Sizer model, i.e., Γ(τ d) and Ω(∆d), can also be obtained from Ω(∆ d) andζ(s b) using probability transformations
Adder model Apart from the free-parameter distributions,ζ(s b) for the Sizer model is obtained from Ω(∆ d) andκ(β) using the arguments presented in Appendix A. The other single-cell-level distributions for the Sizer model, i.e., Γ(τ d) and Ω(∆d), can also be obtained from Ω(∆ d) andζ(s b) using probability transformations. We already have Ω(∆ d) and ζ(s b...
-
[6]
We have sb =βs d, wheres b is the birth-size of the daughter cell ands d is the division-size of the parent cell
Size-related quantities First of all, let us derive the relationships between the mean of the size-related single-cell quantities. We have sb =βs d, wheres b is the birth-size of the daughter cell ands d is the division-size of the parent cell. Taking average on both sides of this equation: ⟨sb⟩=⟨βs d⟩(C1) Since the partitioning fractionβis not correlated...
-
[7]
Division-time for exponential single-cell growth Division-time is a function ofs d,s b, andα.αhere corresponds to the exponential growth rate. For the case of exponential growth, it can be written as: τd = 1 α ln sd sb .(C14) Using Taylor series expansion in the vicinity of the point (⟨s d⟩,⟨s b⟩,⟨α⟩), we can write: τd = 1 ⟨α⟩ ln ⟨sd⟩ ⟨sb⟩ + 1 ⟨α⟩⟨sd⟩(sd ...
-
[8]
Division-time for linear single-cell growth For linear growth, we haveτ d = ∆ d/α, whereαis the linear growth rate. Since, both ∆ d andαare stochastic quantities, we expand the functionτ d about the point(⟨∆ d⟩,⟨α⟩) using Taylor series expansion in two variables: τd = ⟨∆d⟩ ⟨α⟩ + ∞X n=1 (−1)n ⟨∆d⟩ ⟨α⟩ (α− ⟨α⟩) n ⟨α⟩n + ∞X n=0 (−1)n ⟨∆d⟩ ⟨α⟩ ∆d − ⟨∆d⟩ ⟨∆d⟩ ...
-
[9]
Therefore, we have: C(s di , sdj) =⟨s di sdj ⟩ − ⟨sd⟩2 =δ ijσ2 sd ,(D1) wheres di, ands dj are the division-sizes for theith andjth generation cells
Sizer For the Sizer model, division-size of a cell (s d) is selected independently of its birth-size (s b), and the division-sizes of cells from preceding generations in the lineage. Therefore, we have: C(s di , sdj) =⟨s di sdj ⟩ − ⟨sd⟩2 =δ ijσ2 sd ,(D1) wheres di, ands dj are the division-sizes for theith andjth generation cells. And, C(s d, sb) =⟨s dsb⟩...
-
[10]
Therefore, we have: C(∆ di ,∆ dj) =⟨∆ di∆dj ⟩ − ⟨∆d⟩2 =δ ijσ2 ∆d ,(D9) where ∆di, and ∆ dj are the division-added-sizes for theith andjth generation cells
Adder For the Adder model, division-added-size of a cell (∆ d) is selected independently of its birth-size (s b), and the division-added-sizes of cells from preceding generations in the lineage. Therefore, we have: C(∆ di ,∆ dj) =⟨∆ di∆dj ⟩ − ⟨∆d⟩2 =δ ijσ2 ∆d ,(D9) where ∆di, and ∆ dj are the division-added-sizes for theith andjth generation cells. And, C...
-
[11]
2, there are two separate histograms (forζ(s b), Ξ(sd), and Ω(∆d)) for the same experimental data
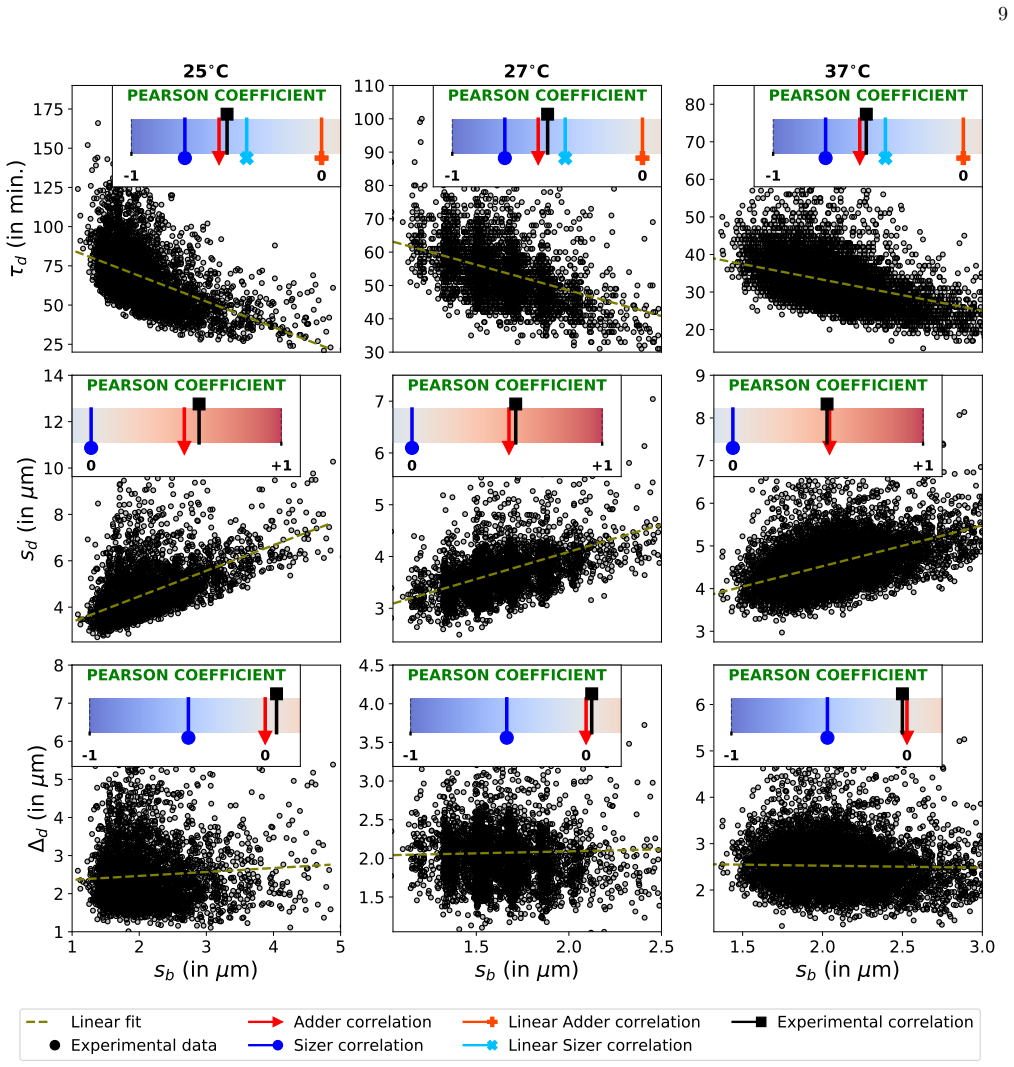
Explanation for two separate histograms for the same experimental data In Fig. 2, there are two separate histograms (forζ(s b), Ξ(sd), and Ω(∆d)) for the same experimental data. The only visible difference is the difference between single-cell growth types. One of them is for exponential growth, whereas the other is for linear growth. The same time-series...
-
[12]
Simulation correlatedα
Extra figures 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 sb (in m) 0.0 0.5 1.0 1.5 2.0 (sb) (A) 2.5 3.0 3.5 4.0 4.5 5.0 5.5 sd (in m) 0.0 0.2 0.4 0.6 0.8 1.0 (sd) (B) 1.0 1.5 2.0 2.5 3.0 3.5 d (in m) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 ( d) (C) 20 30 40 50 60 70 80 90 d (in min.) 0.00 0.01 0.02 0.03 0.04 0.05 ( d) (D) Experimental Exponential Experimental Linear Ana...
2024
-
[13]
Schaechter, O
M. Schaechter, O. Maaløe, and N. O. Kjeldgaard, Dependency on medium and temperature of cell size and chemical composition during balanced growth of salmonella typhimurium, Microbiology19, 592 (1958)
1958
-
[14]
Kjeldgaard, O
N. Kjeldgaard, O. Maaløe, and M. Schaechter, The transition between different physiological states during balanced growth of salmonella typhimurium, Microbiology19, 607 (1958)
1958
-
[15]
Scott, C
M. Scott, C. W. Gunderson, E. M. Mateescu, Z. Zhang, and T. Hwa, Interdependence of cell growth and gene expression: origins and consequences, Science330, 1099 (2010)
2010
-
[16]
Scott, S
M. Scott, S. Klumpp, E. M. Mateescu, and T. Hwa, Emergence of robust growth laws from optimal regulation of ribosome synthesis, Molecular systems biology10, 747 (2014)
2014
-
[17]
W. D. Donachie, Relationship between cell size and time of initiation of dna replication, Nature219, 1077 (1968)
1968
-
[18]
Cooper and C
S. Cooper and C. E. Helmstetter, Chromosome replication and the division cycle of escherichia coli br, Journal of molecular biology31, 519 (1968)
1968
-
[19]
Powell, Growth rate and generation time of bacteria, with special reference to continuous culture, Microbiology15, 492 (1956)
E. Powell, Growth rate and generation time of bacteria, with special reference to continuous culture, Microbiology15, 492 (1956)
1956
-
[20]
Koch and M
A. Koch and M. Schaechter, A model for statistics of the cell division process, Microbiology29, 435 (1962)
1962
-
[21]
Powell, A note on koch & schaechter’s hypothesis about growth and fission of bacteria, Microbiology37, 231 (1964)
E. Powell, A note on koch & schaechter’s hypothesis about growth and fission of bacteria, Microbiology37, 231 (1964)
1964
-
[22]
A. E. Wheals, Size control models of saccharomyces cerevisiae cell proliferation, Molecular and cellular biology2, 361 (1982)
1982
-
[23]
Diekmann, H
O. Diekmann, H. Lauwerier, T. Aldenberg, and J. Metz, Growth, fission and the stable size distribution, Journal of mathematical biology18, 135 (1983)
1983
-
[24]
J. J. Tyson and O. Diekmann, Sloppy size control of the cell division cycle, Journal of theoretical biology118, 405 (1986)
1986
-
[25]
Koppes, M
L. Koppes, M. Meyer, H. Oonk, M. De Jong, and N. Nanninga, Correlation between size and age at different events in the cell division cycle of escherichia coli, Journal of bacteriology143, 1241 (1980)
1980
-
[26]
Voorn and L
W. Voorn and L. Koppes, Skew or third moment of bacterial generation times, Archives of microbiology169, 43 (1997)
1997
-
[27]
Facchetti, F
G. Facchetti, F. Chang, and M. Howard, Controlling cell size through sizer mechanisms, Current Opinion in Systems Biology5, 86 (2017)
2017
-
[28]
Miotto, S
M. Miotto, S. Scalise, M. Leonetti, G. Ruocco, G. Peruzzi, and G. Gosti, A size-dependent division strategy accounts for leukemia cell size heterogeneity, Communications Physics7, 248 (2024)
2024
-
[29]
Nieto, J
C. Nieto, J. Arias-Castro, C. S´ anchez, C. Vargas-Garc´ ıa, and J. M. Pedraza, Unification of cell division control strategies through continuous rate models, Physical Review E101, 022401 (2020)
2020
-
[30]
Santi, N
I. Santi, N. Dhar, D. Bousbaine, Y. Wakamoto, and J. D. McKinney, Single-cell dynamics of the chromosome replication and cell division cycles in mycobacteria, Nature communications4, 2470 (2013)
2013
-
[31]
Campos, I
M. Campos, I. V. Surovtsev, S. Kato, A. Paintdakhi, B. Beltran, S. E. Ebmeier, and C. Jacobs-Wagner, A constant size extension drives bacterial cell size homeostasis, Cell159, 1433 (2014)
2014
-
[32]
Taheri-Araghi, S
S. Taheri-Araghi, S. Bradde, J. T. Sauls, N. S. Hill, P. A. Levin, J. Paulsson, M. Vergassola, and S. Jun, Cell-size control and homeostasis in bacteria, Current biology25, 385 (2015)
2015
-
[33]
Fievet, A
A. Fievet, A. Ducret, T. Mignot, O. Valette, L. Robert, R. Pardoux, A. R. Dolla, and C. Aubert, Single-cell analysis of growth and cell division of the anaerobe desulfovibrio vulgaris hildenborough, Frontiers in microbiology6, 1378 (2015)
2015
-
[34]
Deforet, D
M. Deforet, D. Van Ditmarsch, and J. B. Xavier, Cell-size homeostasis and the incremental rule in a bacterial pathogen, Biophysical journal109, 521 (2015). 33
2015
-
[35]
Priestman, P
M. Priestman, P. Thomas, B. D. Robertson, and V. Shahrezaei, Mycobacteria modify their cell size control under sub- optimal carbon sources, Frontiers in cell and developmental biology5, 64 (2017)
2017
-
[36]
E. S. Chung, P. Kar, M. Kamkaew, A. Amir, and B. B. Aldridge, Single-cell imaging of the mycobacterium tuberculosis cell cycle reveals linear and heterogenous growth, Nature Microbiology , 1 (2024)
2024
-
[37]
J. J. Messelink, F. Meyer, M. Bramkamp, and C. P. Broedersz, Single-cell growth inference of corynebacterium glutamicum reveals asymptotically linear growth, Elife10, e70106 (2021)
2021
-
[38]
M. M. Logsdon, P.-Y. Ho, K. Papavinasasundaram, K. Richardson, M. Cokol, C. M. Sassetti, A. Amir, and B. B. Aldridge, A parallel adder coordinates mycobacterial cell-cycle progression and cell-size homeostasis in the context of asymmetric growth and organization, Current Biology27, 3367 (2017)
2017
-
[39]
Amir, Cell size regulation in bacteria, Physical review letters112, 208102 (2014)
A. Amir, Cell size regulation in bacteria, Physical review letters112, 208102 (2014)
2014
-
[40]
Jun and S
S. Jun and S. Taheri-Araghi, Cell-size maintenance: universal strategy revealed, Trends in microbiology23, 4 (2015)
2015
-
[41]
J. T. Sauls, D. Li, and S. Jun, Adder and a coarse-grained approach to cell size homeostasis in bacteria, Current opinion in cell biology38, 38 (2016)
2016
-
[42]
Chandler-Brown, K
D. Chandler-Brown, K. M. Schmoller, Y. Winetraub, and J. M. Skotheim, The adder phenomenon emerges from indepen- dent control of pre-and post-start phases of the budding yeast cell cycle, Current Biology27, 2774 (2017)
2017
-
[43]
Soifer, L
I. Soifer, L. Robert, and A. Amir, Single-cell analysis of growth in budding yeast and bacteria reveals a common size regulation strategy, Current Biology26, 356 (2016)
2016
-
[44]
Cadart, S
C. Cadart, S. Monnier, J. Grilli, P. J. S´ aez, N. Srivastava, R. Attia, E. Terriac, B. Baum, M. Cosentino-Lagomarsino, and M. Piel, Size control in mammalian cells involves modulation of both growth rate and cell cycle duration, Nature communications9, 3275 (2018)
2018
-
[45]
Willis and K
L. Willis and K. C. Huang, Sizing up the bacterial cell cycle, Nature Reviews Microbiology15, 606 (2017)
2017
-
[46]
Vuaridel-Thurre, A
G. Vuaridel-Thurre, A. R. Vuaridel, N. Dhar, and J. D. McKinney, Computational analysis of the mutual constraints between single-cell growth and division control models, Advanced biosystems4, 1900103 (2020)
2020
-
[47]
Pirie, Being right size, Annual review of microbiology27, 119 (1973)
N. Pirie, Being right size, Annual review of microbiology27, 119 (1973)
1973
-
[48]
W. F. Marshall, K. D. Young, M. Swaffer, E. Wood, P. Nurse, A. Kimura, J. Frankel, J. Wallingford, V. Walbot, X. Qu, et al., What determines cell size?, BMC biology10, 1 (2012)
2012
-
[49]
K. A. Dill, K. Ghosh, and J. D. Schmit, Physical limits of cells and proteomes, Proceedings of the National Academy of Sciences108, 17876 (2011)
2011
-
[50]
A. A. Amodeo and J. M. Skotheim, Cell-size control, Cold Spring Harbor perspectives in biology8, a019083 (2016)
2016
-
[51]
G. E. Neurohr, R. L. Terry, J. Lengefeld, M. Bonney, G. P. Brittingham, F. Moretto, T. P. Miettinen, L. P. Vaites, L. M. Soares, J. A. Paulo,et al., Excessive cell growth causes cytoplasm dilution and contributes to senescence, cell176, 1083 (2019)
2019
-
[52]
Robert, Size sensors in bacteria, cell cycle control, and size control, Frontiers in microbiology6, 515 (2015)
L. Robert, Size sensors in bacteria, cell cycle control, and size control, Frontiers in microbiology6, 515 (2015)
2015
-
[53]
F. Si, G. Le Treut, J. T. Sauls, S. Vadia, P. A. Levin, and S. Jun, Mechanistic origin of cell-size control and homeostasis in bacteria, Current Biology29, 1760 (2019)
2019
-
[54]
P. P. Pandey, H. Singh, and S. Jain, Exponential trajectories, cell size fluctuations, and the adder property in bacteria follow from simple chemical dynamics and division control, Physical Review E101, 062406 (2020)
2020
-
[55]
Nieto, C
C. Nieto, C. A. Vargas-Garcia, and A. Singh, A generalized adder mechanism for cell size homeostasis: Implications for stochastic dynamics of clonal proliferation, bioRxiv (2024)
2024
-
[56]
Iyer-Biswas, C
S. Iyer-Biswas, C. S. Wright, J. T. Henry, K. Lo, S. Burov, Y. Lin, G. E. Crooks, S. Crosson, A. R. Dinner, and N. F. Scherer, Scaling laws governing stochastic growth and division of single bacterial cells, Proceedings of the National Academy of Sciences111, 15912 (2014)
2014
-
[57]
P. Kar, S. Tiruvadi-Krishnan, J. M¨ annik, J. M¨ annik, and A. Amir, Distinguishing different modes of growth using single-cell data, Elife10, e72565 (2021)
2021
-
[58]
Tanouchi, A
Y. Tanouchi, A. Pai, H. Park, S. Huang, N. E. Buchler, and L. You, Long-term growth data of escherichia coli at a single-cell level, Scientific data4, 1 (2017)
2017
-
[59]
P. Wang, L. Robert, J. Pelletier, W. L. Dang, F. Taddei, A. Wright, and S. Jun, Robust growth of escherichia coli, Current biology20, 1099 (2010)
2010
-
[60]
Garc´ ıa-Garc´ ıa, A
R. Garc´ ıa-Garc´ ıa, A. Genthon, and D. Lacoste, Linking lineage and population observables in biological branching processes, Physical Review E99, 042413 (2019)
2019
-
[61]
Genthon and D
A. Genthon and D. Lacoste, Fluctuation relations and fitness landscapes of growing cell populations, Scientific Reports 10, 11889 (2020)
2020
-
[62]
Genthon, Analytical cell size distribution: lineage-population bias and parameter inference, Journal of the Royal Society Interface19, 20220405 (2022)
A. Genthon, Analytical cell size distribution: lineage-population bias and parameter inference, Journal of the Royal Society Interface19, 20220405 (2022)
2022
-
[63]
Donachie, K
W. Donachie, K. Begg, and M. Vicente, Cell length, cell growth and cell division, Nature264, 328 (1976)
1976
-
[64]
Kubitschek, Bilinear cell growth of escherichia coli, Journal of Bacteriology148, 730 (1981)
H. Kubitschek, Bilinear cell growth of escherichia coli, Journal of Bacteriology148, 730 (1981)
1981
-
[65]
J. H. van Heerden, A. Berkvens, D. H. de Groot, and F. J. Bruggeman, Growth consequences of the inhomogeneous organization of the bacterial cytoplasm, BioRxiv , 2023 (2023)
2023
-
[66]
Cylke and S
A. Cylke and S. Banerjee, Super-exponential growth and stochastic size dynamics in rod-like bacteria, Biophysical Journal 122, 1254 (2023)
2023
-
[67]
Osella, E
M. Osella, E. Nugent, and M. Cosentino Lagomarsino, Concerted control of escherichia coli cell division, Proceedings of the National Academy of Sciences111, 3431 (2014)
2014
-
[68]
M. T. Hannebelle, J. X. Ven, C. Toniolo, H. A. Eskandarian, G. Vuaridel-Thurre, J. D. McKinney, and G. E. Fantner, A biphasic growth model for cell pole elongation in mycobacteria, Nature communications11, 452 (2020). 34
2020
-
[69]
Nordholt, J
N. Nordholt, J. H. van Heerden, and F. J. Bruggeman, Biphasic cell-size and growth-rate homeostasis by single bacillus subtilis cells, Current Biology30, 2238 (2020)
2020
-
[70]
Wallden, D
M. Wallden, D. Fange, E. G. Lundius, ¨O. Baltekin, and J. Elf, The synchronization of replication and division cycles in individual e. coli cells, Cell166, 729 (2016)
2016
-
[71]
Serbanescu, N
D. Serbanescu, N. Ojkic, and S. Banerjee, Nutrient-dependent trade-offs between ribosomes and division protein synthesis control bacterial cell size and growth, Cell reports32(2020)
2020
-
[72]
Nieto, C
C. Nieto, C. A. Vargas-Garc´ ıa, J. M. Pedraza, and A. Singh, Mechanisms of cell size regulation in slow-growing escherichia coli cells: Discriminating models beyond the adder, npj Systems Biology and Applications10, 61 (2024)
2024
-
[73]
K. F. Riley, M. P. Hobson, and S. J. Bence,Mathematical Methods for Physics and Engineering: A Comprehensive Guide, 3rd ed. (Cambridge University Press, 2006)
2006
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.