Evaluation of Population Initialization Methods for Genetic Programming-based Symbolic Regression
Pith reviewed 2026-07-01 02:00 UTC · model grok-4.3
The pith
The choice of initialization method has negligible impact on final results in genetic programming for symbolic regression when initial diversity is similar.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
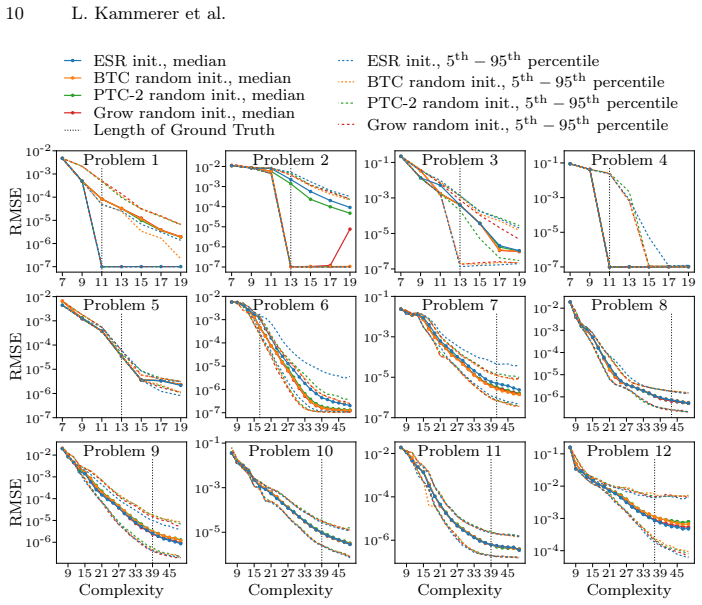
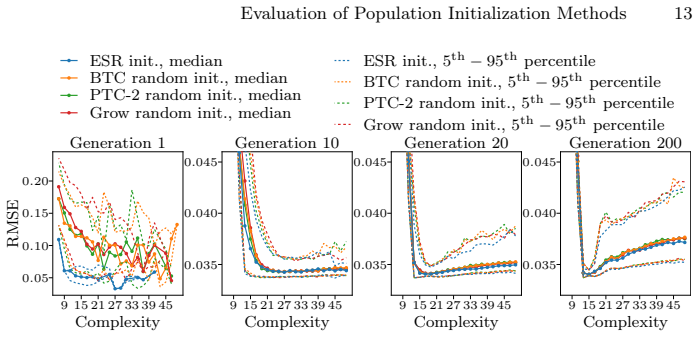
We find no significant differences in accuracy or model complexity among the initialization methods. The initial advantage of initialization with ESR disappears after only a few generations. Our results show that, given similar diversity in the initial population, the effect of the initialization method in GP-based symbolic regression on the final Pareto front is negligible.
What carries the argument
Comparison of Pareto fronts produced by NSGA-II evolutionary search starting from different initial populations in genetic programming symbolic regression.
If this is right
- The initial advantage of using optimized small solutions vanishes quickly during evolution.
- Random initialization methods achieve comparable final accuracy and complexity trade-offs.
- The effect of initialization is negligible when diversity levels are matched across methods.
- Results are consistent across synthetic problems of different complexities and a real-world dataset.
Where Pith is reading between the lines
- Effort in symbolic regression via GP might be better spent on other parameters like selection or variation operators rather than initialization.
- The finding could extend to other multi-objective evolutionary algorithms where population diversity is a key factor.
- Further tests on a wider range of real-world datasets would help establish the generality of the negligible effect.
Load-bearing premise
The twelve synthetic problems and the one real-world dataset are representative of typical symbolic regression tasks.
What would settle it
Demonstrating a symbolic regression problem where one of the initialization methods produces a statistically superior Pareto front after the evolutionary process completes.
Figures







read the original abstract
We analyze the effect of optimizing the initial population of genetic programming (GP) for symbolic regression (SR) on the accuracy and complexity of solutions. We compare three well-established random initialization methods as well as initialization with small optimized solutions from exhaustive symbolic regression (ESR) using a GP/SR implementation which is based on the multi-objective evolutionary algorithm NSGA-II. We compare the final Pareto fronts found with each initialization method on twelve synthetic problems of varying complexity and one real-world dataset. We find no significant differences in accuracy or model complexity among the initialization methods. The initial advantage of initialization with ESR disappears after only a few generations. Our results show that, given similar diversity in the initial population, the effect of the initialization method in GP-based symbolic regression on the final Pareto front is negligible.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper empirically compares three random population initialization methods against initialization with small optimized solutions from exhaustive symbolic regression (ESR) in a multi-objective GP setup based on NSGA-II for symbolic regression. Experiments on twelve synthetic benchmarks of varying complexity and one real-world dataset show no significant differences in final Pareto fronts for accuracy and complexity; any initial ESR advantage vanishes after a few generations. The headline conclusion is that, given comparable initial diversity, the choice of initialization method has negligible impact on GP-SR outcomes.
Significance. If the diversity-equivalence premise and statistical claims hold, the result would indicate that GP-SR performance is robust to standard initialization choices once populations have comparable diversity, reducing the incentive to invest in specialized initialization routines and shifting attention to selection, variation, or objective design. The use of a reproducible multi-objective framework and a mix of synthetic plus real data strengthens the practical relevance of the finding.
major comments (2)
- [Abstract and §4] Abstract and §4 (Results): The central claim is explicitly conditional on 'given similar diversity in the initial population,' yet no diversity metrics (genotypic/phenotypic diversity, tree-size histograms, operator frequencies, or number of unique expressions) are reported at generation 0 to confirm that the ESR initialization produces statistically equivalent diversity to the three random methods. Without this verification the conditional conclusion does not follow from the experiments.
- [§3 and §4] §3 (Experimental Setup) and §4: The abstract asserts 'no significant differences' but the provided text supplies no information on the number of independent runs, the statistical tests employed, correction for multiple comparisons, p-value thresholds, or effect-size reporting. These details are required to evaluate whether the null result is powered and reliable.
minor comments (1)
- [Tables] Table captions and axis labels should explicitly state the diversity measure (if any) used to support the 'similar diversity' premise.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback. We address each major comment below.
read point-by-point responses
-
Referee: [Abstract and §4] Abstract and §4 (Results): The central claim is explicitly conditional on 'given similar diversity in the initial population,' yet no diversity metrics (genotypic/phenotypic diversity, tree-size histograms, operator frequencies, or number of unique expressions) are reported at generation 0 to confirm that the ESR initialization produces statistically equivalent diversity to the three random methods. Without this verification the conditional conclusion does not follow from the experiments.
Authors: We agree that explicit diversity metrics at generation 0 are required to substantiate the conditional claim. The revised manuscript will report phenotypic diversity (number of unique expressions) and tree-size distributions for the initial populations of all methods to allow verification of diversity equivalence. revision: yes
-
Referee: [§3 and §4] §3 (Experimental Setup) and §4: The abstract asserts 'no significant differences' but the provided text supplies no information on the number of independent runs, the statistical tests employed, correction for multiple comparisons, p-value thresholds, or effect-size reporting. These details are required to evaluate whether the null result is powered and reliable.
Authors: We agree that these statistical details are absent from the current text. The revised manuscript will expand §3 to include the number of independent runs, the statistical tests applied, corrections for multiple comparisons, p-value thresholds, and effect-size reporting. revision: yes
Circularity Check
No circularity: purely empirical comparison with no derivations or self-referential steps
full rationale
The paper conducts direct experimental runs of NSGA-II-based GP on 12 synthetic benchmarks plus one real dataset, comparing four initialization methods and reporting final Pareto fronts. No equations, fitted parameters renamed as predictions, uniqueness theorems, or ansatzes appear. The central claim rests on observed outcomes after a few generations rather than any definitional reduction or self-citation chain. The 'given similar diversity' qualifier is an empirical precondition stated in the abstract but does not create circularity because diversity is an observable input property measured (or assumed) before the runs, not derived from the final result.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption NSGA-II is an appropriate algorithm for multi-objective symbolic regression balancing accuracy and complexity.
Reference graph
Works this paper leans on
-
[1]
In: Proceedings of the Genetic and Evolutionary Com- putation Conference Companion
Ahmad, H., Helmuth, T.: A comparison of semantic-based initialization methods for genetic programming. In: Proceedings of the Genetic and Evolutionary Com- putation Conference Companion. pp. 1878–1881 (2018)
2018
-
[2]
Bartlett, D.J., Desmond, H., Ferreira, P.G.: Exhaustive symbolic regression func- tion sets (2022), https://doi.org/10.5281/zenodo.7339113
-
[3]
IEEE Transactions on Evolutionary Computation28(4), 950–964 (2023)
Bartlett, D.J., Desmond, H., Ferreira, P.G.: Exhaustive symbolic regression. IEEE Transactions on Evolutionary Computation28(4), 950–964 (2023)
2023
-
[4]
In: Genetic and Evolutionary Computation Conference
Burke, E., Gustafson, S., Kendall, G.: Ramped half-n-half initialisation bias in gp. In: Genetic and Evolutionary Computation Conference. pp. 1800–1801. Springer (2003)
2003
-
[5]
In: Inter- national Conference on Computer Aided Systems Theory
Burlacu, B., Kammerer, L., Affenzeller, M., Kronberger, G.: Hash-based tree simi- larity and simplification in genetic programming for symbolic regression. In: Inter- national Conference on Computer Aided Systems Theory. pp. 361–369. Springer (2020) Evaluation of Population Initialization Methods 15
2020
-
[6]
In: Proceedings of the 2020 Genetic and Evolutionary Computation Conference Companion
Burlacu, B., Kronberger, G., Kommenda, M.: Operon C++ an efficient genetic pro- gramming framework for symbolic regression. In: Proceedings of the 2020 Genetic and Evolutionary Computation Conference Companion. pp. 1562–1570 (2020)
2020
-
[7]
IEEE transactions on evolutionary computation6(2), 182–197 (2002)
Deb, K., Pratap, A., Agarwal, S., Meyarivan, T.: A fast and elitist multiobjective genetic algorithm: NSGA-II. IEEE transactions on evolutionary computation6(2), 182–197 (2002)
2002
-
[8]
Mathematical problems in engineering2015(1), 212794 (2015)
Deng, Y., Liu, Y., Zhou, D.: An improved genetic algorithm with initial population strategy for symmetric tsp. Mathematical problems in engineering2015(1), 212794 (2015)
2015
-
[9]
Science advances6(5), eaav6971 (2020)
Guimerà, R., Reichardt, I., Aguilar-Mogas, A., Massucci, F.A., Miranda, M., Pal- larès, J., Sales-Pardo, M.: A bayesian machine scientist to aid in the solution of challenging scientific problems. Science advances6(5), eaav6971 (2020)
2020
-
[10]
Information 9(7), 167 (2018)
Hassanat, A.B., Prasath, V.S., Abbadi, M.A., Abu-Qdari, S.A., Faris, H.: An im- proved genetic algorithm with a new initialization mechanism based on regression techniques. Information 9(7), 167 (2018)
2018
-
[11]
MIT Press (1992)
Koza, J.R.: Genetic Programming: On the Programming of Computers by Means of Natural Selection. MIT Press (1992)
1992
-
[12]
CRC Press (2024)
Kronberger, G., Burlacu, B., Kommenda, M., Winkler, S.M., Affenzeller, M.: Sym- bolic regression. CRC Press (2024)
2024
-
[13]
Kronberger, G., de Franca, F.O., Bartlett, D.J., Desmond, H., Ferreira, P.G.: Guid- ingmulti-objectivegeneticprogrammingwithdescriptionlengthimprovessymbolic regression solutions. arXiv preprint arXiv:2605.22374 (2026)
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[14]
In: Proceedings of the Neural Information Processing Systems Track on Datasets and Benchmarks
La Cava, W., Orzechowski, P., Burlacu, B., de França, F.O., Virgolin, M., Jin, Y., Kommenda, M., Moore, J.: Contemporary symbolic regression methods and their relative performance. In: Proceedings of the Neural Information Processing Systems Track on Datasets and Benchmarks. vol. 1 (2021)
2021
-
[15]
In: Euro- pean Conference on Genetic Programming
Langdon, W.B., Nordin, J.: Seeding genetic programming populations. In: Euro- pean Conference on Genetic Programming. pp. 304–315. Springer (2000)
2000
-
[16]
Quarterly of applied mathematics2(2), 164–168 (1944)
Levenberg, K.: A method for the solution of certain non-linear problems in least squares. Quarterly of applied mathematics2(2), 164–168 (1944)
1944
-
[17]
IEEE Trans- actions on Evolutionary Computation4(3), 274–283 (2002)
Luke, S.: Two fast tree-creation algorithms for genetic programming. IEEE Trans- actions on Evolutionary Computation4(3), 274–283 (2002)
2002
-
[18]
Journal of the society for Industrial and Applied Mathematics11(2), 431–441 (1963)
Marquardt, D.W.: An algorithm for least-squares estimation of nonlinear parame- ters. Journal of the society for Industrial and Applied Mathematics11(2), 431–441 (1963)
1963
-
[19]
In: Proceedings of the 35th International Conference on Neural Information Processing Systems
Mundhenk, T.N., Landajuela, M., Glatt, R., Santiago, C.P., Faissol, D.M., Pe- tersen, B.K.: Symbolic regression via neural-guided genetic programming popu- lation seeding. In: Proceedings of the 35th International Conference on Neural Information Processing Systems. pp. 24912–24923 (2021)
2021
-
[20]
VDI-Forschungsheft361, 1 (1933)
Nikuradse, J.: Strömungsgesetze in rauhen Rohren. VDI-Forschungsheft361, 1 (1933)
1933
-
[21]
Journal of Computational Science5(2), 277–297 (2014)
Paul, P.V., Ramalingam, A., Baskaran, R., Dhavachelvan, P., Vivekanandan, K., Subramanian, R.: A new population seeding technique for permutation-coded ge- netic algorithm: Service transfer approach. Journal of Computational Science5(2), 277–297 (2014)
2014
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.