Effect of environmental variation on the benefits of learning
Pith reviewed 2026-06-30 01:05 UTC · model grok-4.3
The pith
Environmental variation distorts egg signals and offsets fitness gains from learning in brood parasitism hosts.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
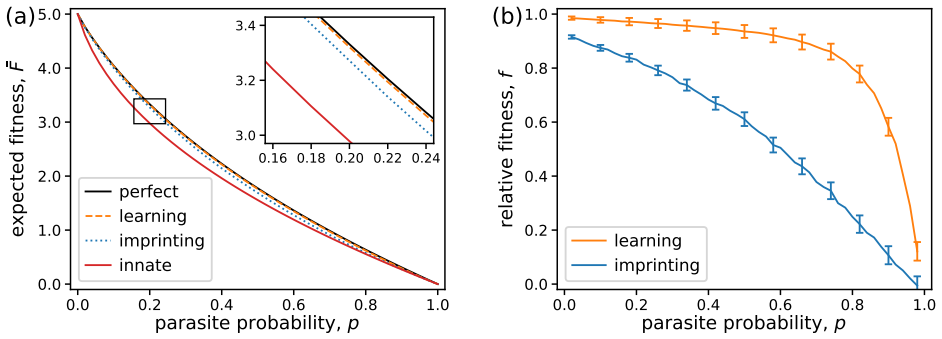
In the signal detection model, phenotypic variation within individuals hinders learning by adding noise to the signals, whereas genotypic variation between individuals promotes learning by carrying more information. Environmental variation that causes egg appearances to fluctuate across clutches reduces the fitness of learning hosts by distorting the signals and creating an effective cost that can offset the benefits of learning, implying that learning or even brief imprinting may be disfavored in homogeneous populations and variable environments.
What carries the argument
Signal detection and learning model incorporating constitutive and induced costs, with sources of variation (phenotypic within hosts, genotypic between hosts, and temporal environmental) acting on egg appearance templates.
If this is right
- Phenotypic variation within a host reduces the value of learning by adding noise.
- Genotypic variation between hosts increases the value of learning by supplying more information.
- Temporal environmental variation imposes an effective cost on learners that can eliminate net fitness gains.
- Learning is less likely to evolve or persist in uniform populations under fluctuating conditions.
Where Pith is reading between the lines
- Similar signal-distortion costs could apply to other learned recognition tasks, such as predator avoidance or mate choice, when environments change rapidly.
- Brief imprinting periods might still be favored if they occur before major environmental shifts, but only if the model is extended to include timing of variation.
- Empirical tests could compare egg-rejection rates in stable versus fluctuating lab or field conditions using genetically similar host lines.
Load-bearing premise
The model treats environmental changes as purely distorting learned templates without any compensating mechanisms that might preserve or restore fitness benefits.
What would settle it
Measure fitness of learning versus non-learning hosts in a genetically uniform population where egg appearances vary over successive clutches; if learners maintain higher fitness, the offset-cost claim does not hold.
Figures








read the original abstract
Signal recognition plays a critical role in species interactions and can be enhanced by learning signal characteristics through experience. In brood parasitism, host species may use visual cues to recognize and reject parasite eggs from their nests; because egg appearances vary within and between host individuals, a host can improve recognition by learning a tailored template of its own eggs. Nevertheless, constitutive and induced costs of learning may inhibit an extended learning period. We use a simple model of signal detection and learning to study how the benefits of learning are affected by different sources of variation in the learning signal. We find that phenotypic variation in egg appearances within a host hinders learning by adding noise to the signals, whereas genotypic variation between individuals promotes learning by carrying more information in the signals. Moreover, we consider environmental variation that can cause egg appearances to fluctuate across clutches over time. We find that such environmental variation reduces the fitness of learning hosts by distorting the signals, creating an effective cost that can offset the benefits of learning. Our results imply that learning or even a brief period of imprinting may be evolutionarily disfavored in homogeneous populations and variable environments.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a simple model of signal detection and learning to examine egg recognition in brood parasitism. Hosts can learn a tailored template of their own eggs to improve rejection of parasite eggs, but learning incurs constitutive and induced costs. The model explores three sources of variation in the learning signal: phenotypic variation within a host (adds noise, hinders learning), genotypic variation between hosts (carries information, promotes learning), and temporal environmental variation across clutches (distorts signals and imposes an effective cost that can offset learning benefits). The central result is that environmental variation can make learning or even brief imprinting evolutionarily disfavored in homogeneous populations.
Significance. If the quantitative results hold under the model's assumptions, the work identifies environmental variation as a previously under-appreciated cost that can reverse the net benefit of learning in recognition systems. This supplies a mechanistic explanation for why learning may be limited in certain ecological settings and offers testable predictions for the evolution of imprinting versus constitutive recognition in host-parasite systems.
major comments (2)
- [Model and environmental-variation results] The claim that environmental variation across clutches creates a net fitness cost sufficient to offset learning benefits depends on the precise implementation of the template-updating rule and the statistical properties of the environmental fluctuations (additive vs. multiplicative, temporal correlation). The manuscript should demonstrate that the reported offset is robust to plausible alternative fluctuation statistics; otherwise the effective-cost conclusion is sensitive to modeling choices that are not shown to be necessary.
- [Methods and Results sections on environmental variation] The quantitative demonstration that environmental variation reduces fitness of learning hosts requires explicit reporting of the updating rule, the form of the signal distributions before and after distortion, the constitutive versus induced cost parameters, and the simulation protocol (including whether hosts may re-sample across multiple clutches). Without these, it is not possible to verify that the distortion alters mean template match rather than merely increasing variance.
minor comments (2)
- [Abstract] The abstract would be strengthened by a single sentence indicating the modeling framework (signal-detection theory with explicit cost terms) and the key equations or simulation approach.
- [Discussion] Clarify in the discussion whether the model allows hosts to average templates over multiple clutches as a potential mitigation of temporal environmental variation.
Simulated Author's Rebuttal
We thank the referee for their thoughtful and constructive review. The comments highlight important aspects of model transparency and robustness that we address below. We have revised the manuscript accordingly to provide the requested details and additional analyses.
read point-by-point responses
-
Referee: [Model and environmental-variation results] The claim that environmental variation across clutches creates a net fitness cost sufficient to offset learning benefits depends on the precise implementation of the template-updating rule and the statistical properties of the environmental fluctuations (additive vs. multiplicative, temporal correlation). The manuscript should demonstrate that the reported offset is robust to plausible alternative fluctuation statistics; otherwise the effective-cost conclusion is sensitive to modeling choices that are not shown to be necessary.
Authors: We agree that robustness to modeling choices is important for the strength of the conclusion. In the revised manuscript we have added supplementary analyses demonstrating that the effective cost persists under both additive and multiplicative environmental fluctuations and across a range of temporal correlation strengths. The qualitative result that environmental variation can offset learning benefits remains consistent, although the precise magnitude of the offset varies with the fluctuation statistics as expected. revision: yes
-
Referee: [Methods and Results sections on environmental variation] The quantitative demonstration that environmental variation reduces fitness of learning hosts requires explicit reporting of the updating rule, the form of the signal distributions before and after distortion, the constitutive versus induced cost parameters, and the simulation protocol (including whether hosts may re-sample across multiple clutches). Without these, it is not possible to verify that the distortion alters mean template match rather than merely increasing variance.
Authors: We accept this criticism and have substantially expanded the Methods section. The revised text now explicitly states the template-updating rule (incremental Bayesian updating of the mean and variance of the internal template), the pre- and post-distortion signal distributions (Gaussian with environment-induced mean shift), the constitutive and induced cost parameters, and the simulation protocol (single clutch sampled per generation with no re-sampling across clutches). These additions confirm that the primary effect is a systematic shift in the mean template match rather than a pure increase in variance. revision: yes
Circularity Check
No circularity; forward model derives effects from explicit assumptions
full rationale
The paper constructs a signal-detection model with explicit constitutive/induced costs and three distinct variation sources (phenotypic within-host noise, genotypic between-host information, temporal environmental distortion). Fitness outcomes for learning vs. non-learning hosts are computed directly from these inputs via the model's updating rule and detection thresholds. No equation reduces to a fitted parameter or self-referential definition, no uniqueness theorem is imported via self-citation, and no prediction is statistically forced by construction. The derivation chain is therefore self-contained against the stated premises.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Learning has constitutive and induced costs that inhibit an extended learning period.
- domain assumption Signal recognition is enhanced by learning a tailored template from experience in a signal detection framework.
Reference graph
Works this paper leans on
-
[1]
G. D. Ruxton et al.Avoiding attack: the evolutionary ecology of crypsis, aposematism, and mimicry. Oxford university press, 2019. 18
2019
-
[2]
Noisy communities and signal detection: why do foragers visit rewardless flowers?
E. M. Lichtenberg et al. “Noisy communities and signal detection: why do foragers visit rewardless flowers?”Philosophical Transactions of the Royal Society B: Biological Sciences 375.1802 (2020)
2020
-
[3]
Signal categorization by foraging animals depends on ecological diver- sity
D. W. Kikuchi et al. “Signal categorization by foraging animals depends on ecological diver- sity”.Elife8 (2019), e43965
2019
-
[4]
S. J. Shettleworth.Cognition, Evolution, and Behavior. Oxford university press, 2009
2009
-
[5]
Evolutionary biology of animal cognition
R. Dukas. “Evolutionary biology of animal cognition”.Annu. Rev. Ecol. Evol. Syst.35.1 (2004), pp. 347–374
2004
-
[6]
What are the costs of learning? Modest trade-offs and constitutive costs do not set the price of fast associative learning ability in a parasitoid wasp
M. Liefting et al. “What are the costs of learning? Modest trade-offs and constitutive costs do not set the price of fast associative learning ability in a parasitoid wasp”.Animal cognition 22.5 (2019), pp. 851–861
2019
-
[7]
Selective costs and benefits in the evolution of learning
T. D. Johnston. “Selective costs and benefits in the evolution of learning”. In:Advances in the Study of Behavior. Vol. 12. Elsevier, 1982, pp. 65–106
1982
-
[8]
Costs of memory: lessons from ‘mini’brains
J. G. Burns, J. Foucaud, and F. Mery. “Costs of memory: lessons from ‘mini’brains”.Pro- ceedings of the Royal Society B: Biological Sciences278.1707 (2011), pp. 923–929
2011
-
[9]
Reproductive tradeoffs of learning in a butterfly
E. C. Snell-Rood, G. Davidowitz, and D. R. Papaj. “Reproductive tradeoffs of learning in a butterfly”.Behavioral Ecology22.2 (2011), pp. 291–302
2011
-
[10]
Artificial selection on relative brain size in the guppy reveals costs and benefits of evolving a larger brain
A. Kotrschal et al. “Artificial selection on relative brain size in the guppy reveals costs and benefits of evolving a larger brain”.Current Biology23.2 (2013), pp. 168–171
2013
-
[11]
Energetic cost of learning and memory can cause cognitive impairment in honeybees
S. Jaumann, R. Scudelari, and D. Naug. “Energetic cost of learning and memory can cause cognitive impairment in honeybees”.Biology Letters9.4 (2013), p. 20130149
2013
-
[12]
To favor survival under food shortage, the brain disables costly memory
P.-Y. Placais and T. Preat. “To favor survival under food shortage, the brain disables costly memory”.Science339.6118 (2013), pp. 440–442
2013
-
[13]
Costs of learning and the evolution of mimetic signals
D. W. Kikuchi and T. N. Sherratt. “Costs of learning and the evolution of mimetic signals”. American Naturalist186.3 (Sept. 2015), pp. 321–332
2015
-
[14]
Change, regularity, and value in the evolution of animal learning
D. W. Stephens. “Change, regularity, and value in the evolution of animal learning”.Behav- ioral Ecology2.1 (1991), pp. 77–89
1991
-
[15]
Reliability, uncertainty, and costs in the evolution of animal learning
A. S. Dunlap and D. W. Stephens. “Reliability, uncertainty, and costs in the evolution of animal learning”.Current opinion in behavioral sciences12 (2016), pp. 73–79
2016
-
[16]
Mechanisms of avian egg recognition: possible learned and innate factors
S. I. Rothstein. “Mechanisms of avian egg recognition: possible learned and innate factors”. The Auk91.4 (1974), pp. 796–807
1974
-
[17]
Components of change in the evolution of learning and unlearned preference
A. S. Dunlap and D. W. Stephens. “Components of change in the evolution of learning and unlearned preference”.Proceedings of the Royal Society B: Biological Sciences276.1670 (2009), pp. 3201–3208
2009
-
[18]
How do sensitive periods arise and what are they for?
P. Bateson. “How do sensitive periods arise and what are they for?”Animal Behaviour27 (1979), pp. 470–486
1979
-
[19]
Mechanisms of avian egg-recognition: additional evidence for learned com- ponents
S. I. Rothstein. “Mechanisms of avian egg-recognition: additional evidence for learned com- ponents”.Animal Behaviour26 (1978), pp. 671–677
1978
-
[20]
The sensitive period for auditory-vocal learning in the zebra finch: Consequences of limited-model availability and multiple-tutor paradigms on song imitation
S. M. Gobes, R. B. Jennings, and R. K. Maeda. “The sensitive period for auditory-vocal learning in the zebra finch: Consequences of limited-model availability and multiple-tutor paradigms on song imitation”.Behavioural Processes163 (2019), pp. 5–12. 19
2019
-
[21]
Early learning influences species assortative mating pref- erences in Lake Victoria cichlid fish
M. N. Verzijden and C. ten Cate. “Early learning influences species assortative mating pref- erences in Lake Victoria cichlid fish”.Biology letters3.2 (2007), pp. 134–136
2007
-
[22]
Constraints on egg discrimination and cuckoo-host co-evolution
A. Lotem, H. Nakamura, and A. Zahavi. “Constraints on egg discrimination and cuckoo-host co-evolution”.Animal behaviour49.5 (1995), pp. 1185–1209
1995
-
[23]
How to learn to recognize conspecific brood parasitic offspring
D. Shizuka and B. E. Lyon. “How to learn to recognize conspecific brood parasitic offspring”. Philosophical Transactions of the Royal Society B375.1802 (2020), p. 20190472
2020
-
[24]
Exploration or exploitation: life expectancy changes the value of learning in foraging strategies
S. Eliassen et al. “Exploration or exploitation: life expectancy changes the value of learning in foraging strategies”.Oikos116.3 (2007), pp. 513–523
2007
-
[25]
A minimal model of learning: quantifying the cost and benefit of learning in changing environments
E. Arehart and F. R. Adler. “A minimal model of learning: quantifying the cost and benefit of learning in changing environments”.Proceedings of the Royal Society B290.2005 (2023), p. 20231084
2005
-
[26]
How precise is egg discrimination in weaverbirds?
D. C. Lahti and A. R. Lahti. “How precise is egg discrimination in weaverbirds?”Animal Behaviour63.6 (2002), pp. 1135–1142
2002
-
[27]
Foraging errors play a role in resource exploration by bumble bees (Bombus terrrestris)
L. J. Evans and N. E. Raine. “Foraging errors play a role in resource exploration by bumble bees (Bombus terrrestris)”.Journal of Comparative Physiology A200.6 (2014), pp. 475–484
2014
-
[28]
Active defence mechanisms in brood parasitism hosts and their consequences for parasite adaptation and speciation
G. Dillenseger. “Active defence mechanisms in brood parasitism hosts and their consequences for parasite adaptation and speciation”.Journal of Avian Biology2024.11-12 (2024), e03252
2024
-
[29]
Learning to recognize nestlings is maladaptive for cuckoo Cuculus canorus hosts
A. Lotem. “Learning to recognize nestlings is maladaptive for cuckoo Cuculus canorus hosts”. Nature362.6422 (1993), pp. 743–745
1993
-
[30]
Do first-time breeding females imprint on their own eggs?
M. Soler et al. “Do first-time breeding females imprint on their own eggs?”Proceedings of the Royal Society B: Biological Sciences280.1752 (2013), p. 20122518
2013
-
[31]
Parental-care parasitism: how do unrelated offspring attain accep- tance by foster parents?
M. Roldan and M. Soler. “Parental-care parasitism: how do unrelated offspring attain accep- tance by foster parents?”Behavioral Ecology22.4 (2011), pp. 679–691
2011
-
[32]
Bird brood parasitism
M. Stevens. “Bird brood parasitism”.Current Biology23.20 (2013), R909–R913
2013
-
[33]
A brood parasitic catfish of mouthbrooding cichlid fishes in Lake Tanganyika
T. Sato. “A brood parasitic catfish of mouthbrooding cichlid fishes in Lake Tanganyika”. Nature323.6083 (1986), pp. 58–59
1986
-
[34]
Cuckoos versus hosts in insects and birds: adaptations, counter-adaptations and outcomes
R. M. Kilner and N. E. Langmore. “Cuckoos versus hosts in insects and birds: adaptations, counter-adaptations and outcomes”.Biological Reviews86.4 (2011), pp. 836–852
2011
-
[35]
N. B. Davies.Cuckoos, Cowbirds and Other Cheats. T & A D Poyser, 2000
2000
-
[36]
Intraspecific nest parasitism in birds
Y. Yom-Tov. “Intraspecific nest parasitism in birds”.Biological Reviews55.1 (1980), pp. 93– 108
1980
-
[37]
The evolution of virulence in brood parasites
R. M. Kilner. “The evolution of virulence in brood parasites”.Ornithological Science4.1 (2005), pp. 55–64
2005
-
[38]
Cuckoos versus reed warblers: adaptations and counter- adaptations
N. B. Davies and M. d. L. Brooke. “Cuckoos versus reed warblers: adaptations and counter- adaptations”.Animal behaviour36.1 (1988), pp. 262–284
1988
-
[39]
Mechanisms of avian egg recognition: Which egg parameters elicit responses by rejecter species?
S. I. Rothstein. “Mechanisms of avian egg recognition: Which egg parameters elicit responses by rejecter species?”Behavioral Ecology and Sociobiology11.4 (1982), pp. 229–239
1982
-
[40]
Recognizing odd smells and ejection of brood parasitic eggs. An experimental test in magpies of a novel defensive trait against brood parasitism
J. Soler et al. “Recognizing odd smells and ejection of brood parasitic eggs. An experimental test in magpies of a novel defensive trait against brood parasitism”.Journal of Evolutionary Biology27.6 (2014), pp. 1265–1270. 20
2014
-
[41]
A review of the cues used for rejecting foreign eggs from the nest by the Eurasian blackbird (Turdus merula)
A. G. Fulmer and M. E. Hauber. “A review of the cues used for rejecting foreign eggs from the nest by the Eurasian blackbird (Turdus merula)”.Ecology and Evolution12.5 (2022), e8886
2022
-
[42]
Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs
C. N. Spottiswoode and M. Stevens. “Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs”.Proceedings of the National Academy of Sciences107.19 (2010), pp. 8672–8676
2010
-
[43]
Mechanisms of avian egg-recognition: Do birds know their own eggs?
S. I. Rothstein. “Mechanisms of avian egg-recognition: Do birds know their own eggs?”Animal Behaviour23.2 (1975), pp. 268–278
1975
-
[44]
Recognition errors and probability of parasitism determine whether reed warblers should accept or reject mimetic cuckoo eggs
N. B. Davies, M. d. L. Brooke, and A. Kacelnik. “Recognition errors and probability of parasitism determine whether reed warblers should accept or reject mimetic cuckoo eggs”. Proceedings of the Royal Society of London. Series B: Biological Sciences263.1372 (1996), pp. 925–931
1996
-
[45]
S. E. Solomon.Egg and Eggshell Quality. London: Wolfe Publishing, 1991
1991
-
[46]
Environmental conditions influence egg color of reed warblers Acro- cephalus scirpaceus and their parasite, the common cuckoo Cuculus canorus
J. M. Aviles et al. “Environmental conditions influence egg color of reed warblers Acro- cephalus scirpaceus and their parasite, the common cuckoo Cuculus canorus”.Behavioral Ecology and Sociobiology61.3 (2007), pp. 475–485
2007
-
[47]
Effects of environmental conditions on the egg mass, yolk antioxidant level, eggshell thickness and eggshell spotting patterns of Great Tits (Parus major)
R. Hargitai et al. “Effects of environmental conditions on the egg mass, yolk antioxidant level, eggshell thickness and eggshell spotting patterns of Great Tits (Parus major)”.Journal of Ornithology157.4 (2016), pp. 995–1006
2016
-
[48]
How to detect a cuckoo egg: a signal-detection theory model for recognition and learning
M. A. Rodriguez-Girones and A. Lotem. “How to detect a cuckoo egg: a signal-detection theory model for recognition and learning”.The American Naturalist153.6 (1999), pp. 633– 648
1999
-
[49]
M. J. Hautus, N. A. Macmillan, and C. D. Creelman.Detection theory: A user’s guide. Routledge, 2021
2021
-
[50]
Multiparasitism: why do interspecific brood parasites lay eggs so frequently in already parasitized host nests?
M. Soler. “Multiparasitism: why do interspecific brood parasites lay eggs so frequently in already parasitized host nests?”Biological Reviews100.4 (2025), pp. 1660–1679
2025
-
[51]
T. M. Cover and J. A. Thomas.Elements of Information Theory. Wiley, 2005
2005
-
[52]
Optimizing reproduction in a randomly varying environment when a correlation may exist between the conditions at the time a choice has to be made and the subsequent outcome
D. Cohen. “Optimizing reproduction in a randomly varying environment when a correlation may exist between the conditions at the time a choice has to be made and the subsequent outcome”.Journal of Theoretical Biology16.1 (1967), pp. 1–14
1967
-
[53]
The fitness value of infor- mation
M. C. Donaldson-Matasci, C. T. Bergstrom, and M. Lachmann. “The fitness value of infor- mation”.Oikos119.2 (2010), pp. 219–230
2010
-
[54]
The value of information for populations in varying environments
O. Rivoire and S. Leibler. “The value of information for populations in varying environments”. Journal of Statistical Physics142.6 (2011), pp. 1124–1166
2011
-
[55]
Bialek.Biophysics: searching for principles
W. Bialek.Biophysics: searching for principles. Princeton University Press, 2012
2012
-
[56]
Avian vision and the evolution of egg color mimicry in the common cuckoo
M. C. Stoddard and M. Stevens. “Avian vision and the evolution of egg color mimicry in the common cuckoo”.Evolution65.7 (2011), pp. 2004–2013
2011
-
[57]
Age-Related Brood Parasitism and Egg Rejection in Magpie Hosts
J. G. Mart´ ınez et al. “Age-Related Brood Parasitism and Egg Rejection in Magpie Hosts”. The American Naturalist195.5 (2020), pp. 876–885
2020
-
[58]
Lifetime fitness of short-and long-distance dis- persing great reed warblers
B. Hansson, S. Bensch, and D. Hasselquist. “Lifetime fitness of short-and long-distance dis- persing great reed warblers”.Evolution58.11 (2004), pp. 2546–2557. 21
2004
-
[59]
Egg color variation, but not egg rejection behavior, changes in a cuckoo host breeding in the absence of brood parasitism
C. Yang et al. “Egg color variation, but not egg rejection behavior, changes in a cuckoo host breeding in the absence of brood parasitism”.Ecology and Evolution4.11 (2014), pp. 2239– 2246
2014
-
[60]
Genetic and geographic variation in rejection behavior of cuckoo eggs by European magpie populations: An experimental test of rejecter-gene flow
J. J. Soler et al. “Genetic and geographic variation in rejection behavior of cuckoo eggs by European magpie populations: An experimental test of rejecter-gene flow”.Evolution53.3 (1999), pp. 947–956
1999
-
[61]
Do weather conditions affect the colouration of great reed warbler Acrocephalus arundinaceus eggs?
M. Honza, P. Prochazka, and M. Pozgayova. “Do weather conditions affect the colouration of great reed warbler Acrocephalus arundinaceus eggs?”Folia zoologica61.3–4 (2012), pp. 219– 224
2012
-
[62]
A review of quorum-sensing and its role in mediating interkingdom interactions in the ocean
M. Coolahan and K. E. Whalen. “A review of quorum-sensing and its role in mediating interkingdom interactions in the ocean”.Communications Biology8.1 (2025), p. 179
2025
-
[63]
Adaptation and memory in immune responses
G. Natoli and R. Ostuni. “Adaptation and memory in immune responses”.Nature immunology 20.7 (2019), pp. 783–792
2019
-
[64]
Innate and adaptive immune memory: an evolutionary continuum in the host’s response to pathogens
M. G. Netea et al. “Innate and adaptive immune memory: an evolutionary continuum in the host’s response to pathogens”.Cell host & microbe25.1 (2019), pp. 13–26
2019
-
[65]
An empirical test of 2-dimensional signal detection theory applied to Batesian mimicry
D. W. Kikuchi et al. “An empirical test of 2-dimensional signal detection theory applied to Batesian mimicry”.Behavioral Ecology26.4 (2015), pp. 1226–1235
2015
-
[66]
The Companion in the Bird’s World
K. Z. Lorenz. “The Companion in the Bird’s World”.The Auk54.3 (1937), pp. 245–273
1937
-
[67]
Homing in Pacific Salmon: Mechanisms and Ecological Basis
A. H. Dittman and T. P. Quinn. “Homing in Pacific Salmon: Mechanisms and Ecological Basis”.Journal of Experimental Biology199.1 (1996), pp. 83–91
1996
-
[68]
Nongenetic Inheritance and Its Evolutionary Implications
R. Bonduriansky and T. Day. “Nongenetic Inheritance and Its Evolutionary Implications”. Annual Review of Ecology, Evolution, and Systematics40 (2009), pp. 103–125
2009
-
[69]
Transgenerational Epigenetic Inheritance: Myths and Mechanisms
E. Heard and R. A. Martienssen. “Transgenerational Epigenetic Inheritance: Myths and Mechanisms”.Cell157.1 (2014), pp. 95–109
2014
-
[70]
D. M. Green and J. A. Swets.Signal detection theory and psychophysics. Wiley, 1966
1966
-
[71]
Signal detection, acceptance thresholds and the evolution of animal recognition systems
A. V. Suarez et al. “Signal detection, acceptance thresholds and the evolution of animal recognition systems”.Philosophical Transactions of the Royal Society B: Biological Sciences 375.1802 (2020), p. 20190464
2020
-
[72]
Signal detection and optimal acceptance thresholds in avian brood parasite-host systems: implications for egg rejection
F. Ruiz-Raya and M. Soler. “Signal detection and optimal acceptance thresholds in avian brood parasite-host systems: implications for egg rejection”.Philosophical Transactions of the Royal Society B: Biological Sciences375.1802 (2020), p. 20190477. 22
2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.